Visual Neglect
Visual neglect is
a common neurological syndrome in which patients fail to acknowledge stimuli
toward the side of space opposite to their unilateral lesion. This disability
affects many aspects of their life. For example, after a right lesion, patients
typically fail to eat the food located on the left side of their plate, or to
shave or make up the left side of their face, and, in extreme cases, may no
longer acknowledge the left side of their body as their own. In a common
clinical test known as line cancellation, they fail to mark lines toward the
left of the page (see figure 1A) despite often being able to detect isolated
lines presented in their left visual field, demonstrating that they are not
simply blind on that side.
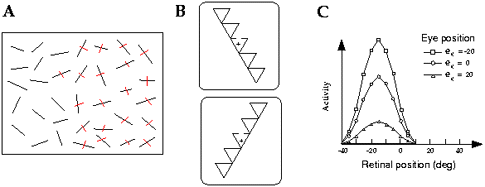
Figure 1 A. Left items neglected in a cancellation
task. B. Visual displays from the Driver et al. (1994) experiment. The cross
indicates the fixation point. Left-neglect patients performed better for the
bottom configuration than the top one, even though the gap to be detected was
at the same retinal location. This pattern is consistent with left
object-centered neglect. C. Visual receptive field of a typical monkey parietal
neuron, for three different eye positions. The retinotopic position of the
receptive field is invariant across eye positions but the gain of the response
changes. (Adapted from Andersen, Issick, and Siegel 1985.)
This syndrome is observed
primarily after unilateral lesions to the parieto-occipital junction,
especially in the right hemisphere (Heilman, Watson, and Valenstein 1985;
Bisiach 1996). Lesions to the right frontal cortex and to various subcortical
sites can also trigger neglect-like symptoms, although with subtle differences
from parietal neglect (Heilman et al. 1985; Guariglia et al. 1993). Here we
concentrate on parietal neglect, as it is the most common form, and can be
related to recent data on the parietal lobe from nonhuman primates.
Two major accounts have been
proposed for neglect. Some theories posit a deficit in directing attention
toward contralesional events (Posner et al. 1984; Kinsbourne 1987; see ATTENTION and ATTENTION IN THE HUMAN
BRAIN). For instance, right parietal patients -- who suffer from left
neglect -- tend to have particular difficulty in detecting stimuli in the left
hemifield if their attention has previously been drawn to the right side
(Posner et al. 1984). By contrast, many "preattentive" aspects of vision
appear to be spared on the affected side (Driver, Baylis, and Rafal 1992;
McGlinchey-Berroth et al. 1996; Mattingley, Davis, and Driver 1997).
Other accounts argue that the
patient's lesion simply disrupts the neuronal coding of contralesional space,
at relatively high levels of representation (Bisiach and Luzzatti 1978;
Bisiach, Luzzati, and Perani 1979; Rizzolatti and Berti 1990; Halligan and
Marshall 1991; Karnath, Schenkel, and Fischer 1991). This perspective has drawn
support from the finding that even mental IMAGERY can be
impaired in some left-neglect patients (Bisiach and Luzzatti 1978), such that
they fail to report what would appear on their left when retrieving from memory
the view of a familiar visual scene.
The dichotomy between
attentional and representational accounts has recently been challenged by
several authors using neural network models in which attentional and
representational functions are interwoven (Mozer and Behrmann 1990; Cohen et
al. 1994; Pouget and Sejnowski 1997a). This work suggests a compromise view,
whereby neglect results from damage to cortical areas that are located at the
interface between sensory and motor systems, and which are responsible for both
the representation of the position of objects and the selective control of
spatial action, that is to say, "attention."
Frames of reference: In
principle, "left" neglect might refer to the left of the visual field,
or the left of the head, or the trunk, or even of the surrounding environment.
To determine the frame of reference for hemineglect, one can test patients in
various postures, so that a stimulus location changes in one frame of reference
while remaining constant in the others. For instance, one might test a patient
looking straight ahead vs. with the gaze deviated twenty degrees to the right,
while keeping all stimuli at the same position with respect to the RETINA. If
neglect were purely retinotopic, these conditions should not differ, whereas if
it were head- or body-centered, performance should change accordingly. Such
experiments have typically revealed that neglect affects a mixture of frames of
reference concurrently, rather than just one single frame. Thus, the
probability that a patient will neglect a particular visual stimulus is
typically a function of its position in various egocentric frames of reference,
such as eye-, head- or trunk-centered, as well as showing influences from cues
in the environment, for example, as regards the gravitational upright (Bisiach,
Capitani, and Porta 1985; Ladavas 1987; Ladavas, Pesce, and Provincial, 1989;
Calvanio, Petrone, and Levine 1987; Farah et al. 1990; Karnath et al. 1991;
Behrmann and Moscovitch 1994).
A few experiments suggest that
visual neglect can also be "object-centered," that is, patients tend
to neglect the left side of an object regardless of its position or orientation
(Driver et al. 1994; Tipper and Behrmann 1996). For example, Driver et al.
(1994) devised a situation in which left-neglect patients could detect a gap in
part of a triangle when this gap was perceived to be on the right side of an
object, but missed the same gap when it was seen as belonging to the left side,
even though it still fell at the same location relative to the patient (figure
1B). Such results seem consistent with the existence of object-centered
representations in the parietal cortex.
Many other studies claim to have
found evidence for object-centered neglect (Driver and Halligan 1991; Arguin
and Bub 1993; Halligan and Marshall 1994), but as pointed out by Driver et al.
(1994), their results could be explained instead by what we will call relative
neglect in strictly egocentric coordinates (see also Kinsbourne 1987; Mozer and
Behrmann 1990; Desimone and Duncan 1995; and Pouget and Sejnowski 1997a for
variations on this idea). When confronted with two competing objects, patients
may neglect the one farther to the left even if both fall in the right
hemispace egocentrically, and likewise for the subparts of a single object
(Driver and Halligan 1991; Driver et al. 1992; Driver et al. 1994; Halligan and
Marshall 1994). Thus, it appears that the relative position of objects
or their subparts is just as important as their absolute position with
respect to the patient. This phenomenon can be explained if the lesion induces
a gradient of neglect with increasing severity in the egocentric
contralesional direction (Kinsbourne 1987; Driver et al. 1994; Pouget and
Sejnowski 1997a).
Neural basis: There have been
several attempts to relate neglect to what is known of the response properties
of parietal neurons from single-cell recordings in monkeys (Mozer and Behrmann
1990; Duhamel et al. 1992; Anderson 1996; Mozer, Halligan, and Marshall 1997;
Pouget and Sejnowski 1997a; see also MODELING
NEUROPSYCHOLOGICAL DEFICITS and SPATIAL PERCEPTION).
Such models generally rely on cells in the parietal cortex having retinotopic
receptive fields, with each hemisphere tending to overrepresent the
contralateral visual field (see, however, Duhamel et al. 1992 for a different
approach). Consequently, a right lesion leads to a neuronal gradient in which
the left side of the retina is less strongly represented than the right side,
producing left neglect. In such models, there is no particular dividing midline
such that any stimulus to the left of it is invariably neglected. Instead,
neglect depends only on the relative position of competing stimuli, as
discussed above, with objects or object parts that are farther toward the
retinal left than their competitors being neglected. These models readily
capture the behavior of patients in tasks such as line bisection, line
cancellation, and in some of the paradigms discussed above that have revealed
relative neglect.
Parietal neurons, however, do
not simply respond to visual stimulation, but also integrate sensory responses
with posture signals such as eye and head position. Andersen and colleagues
have shown that the retinotopic receptive fields of parietal cells are gain-modulated
by such posture signals (Andersen, Essick, and Siegel 1985; Andersen et al.
1997; see figure 1C for an example in which the visual receptive field of a
cell is modulated by eye position). These response properties can be modeled as
basis functions of the inputs, a type of function which is
particularly well-suited to the computational demand of sensorimotor
transformations (Pouget and Sejnowski 1997b).
A simulated unilateral lesion in
such a basis-function representation produces an impairment that resembles
clinical neglect, in that the deficit affects a mixture of egocentric frames of
reference as found in patients (Pouget and Sejnowski 1997a). This approach can
also be generalized to encompass object-centered neglect, as in the Driver et
al. (1994) experiment depicted in figure 1B, by considering the perceived
orientation of the object as providing a signal analogous to the posture
signals integrated by the basis functions (Deneve and Pouget in press). This
basis-function framework can explain why neglect may be influenced by stimulus
position relative to the retina, head, body, other objects, and other parts of
the same object, all at the same time, without requiring cells in the parietal
cortex to have visual receptive fields explicitly defined in any single one of
these frames of reference.
Neglect remains a fascinating
but disabling disorder, which still poses a major challenge to rehabilitation.
Its further study will hopefully lead to more effective treatments, as well as
reveal more about how the brain represents space, and allows for selective
spatial attention.
See also
Additional links
References and Further Readings
Anderson, B.
(1996). A mathematical model of line bisection behaviour in neglect. Brain
119:841-850.
Andersen, R. A.,
G. K. Essick, and R. M. Siegel. (1985). Encoding of spatial location by
posterior parietal neurons. Science 230:456-458.
Andersen, R. A.,
L. H. Snyder, D. C. Bradley, and J . Xing. (1997). Encoding of intention and
spatial location in the posterior parietal cortex. Ann. Rev. Neurosci.
20:303-330.
Arguin, M., and D.
N. Bub. (1993). Evidence for an independent stimulus-centered reference frame
from a case of visual hemi-neglect. Cortex 29:349-357.
Behrmann, M., and
M. Moscovitch. (1994). Object-centered neglect in patients with unilateral
neglect: Effects of left-right coordinates of objects. Journal of
Cognitive Neuroscience 6(2):151-155.
Bisiach, E.
(1996). Unilateral neglect and the structure of space representation. Current
Directions in Psychological Science 5(2):62-65.
Bisiach, E., E.
Capitani, and E. Porta. (1985). Two basic properties of space representation in
the brain: Evidence from unilateral neglect. Journal of Neurology,
Neurosurgery and Psychiatry 48:141-144.
Bisiach, E., and
C. Luzzatti. (1978). Unilateral neglect of representational space. Cortex
14:129-133.
Bisiach, E., C.
Luzzatti, and D. Perani. (1979). Unilateral neglect, representational schema
and consciousness. Brain 102:609-618.
Calvanio, R., P.
N. Petrone, and D. N. Levine. (1987). Left visual spatial neglect is both
environment-centered and body-centered. Neurology 37:1179-1181.
Cohen, J. D., M.
J. Farah, R. D. Romero, and D. Servan-Schreiber. (1994). Mechanisms of spatial
attention: The relation of macrostructure to microstructure in parietal
neglect. Journal of Cognitive Neuroscience 6(4):377-387.
Deneve, S., and A.
Pouget. (Forthcoming). Neural basis of object-centered representations. In Advances
in Neural Information Processing Systems, vol. 11. Cambridge, MA: MIT
Press.
Desimone, R., and
J. Duncan. (1995). Neural mechanisms of selective visual attention. Ann.
Rev. Neurosci. 18:193-222.
Driver, J., and P.
W. Halligan. (1991). Can visual neglect operate in object-centered coordinates?
An affirmative single case study. Cognitive Neuropsychology
8(6):475-496.
Driver, J., G. C.
Baylis, and R. D. Rafal. (1992). Preserved figure-ground segregation and symmetry
perception in visual neglect. Nature 360:73-75.
Driver, J., G. C.
Baylis, S. J. Goodrich, and R. D. Rafal. (1994). Axis-based neglect of visual
shapes. Neuropsychologia 32(11):1353-1365.
Duhamel, J. R., M.
E. Goldberg, E. J. Fitzgibbon, A. Sirigu, and J. Grafman. (1992). Saccadic
dysmetria in a patient with a right frontoparietal lesion. The importance of
corollary discharge for accurate spatial behaviour. Brain
115:1387-1402.
Farah, M. J., J.
L. Brunn, A. B. Wong, M. A. Wallace, and P. A. Carpenter. (1990). Frames of
reference for allocating attention to space: Evidence from the neglect
syndrome. Neuropsychologia 28(4):335-347.
Guariglia, C., A.
Padovani, P. Pantano, and L. Pizzamiglio. (1993). Unilateral neglect restricted
to visual imagery. Nature 364:235-237.
Halligan, P. W.,
and J. C. Marshall. (1991). Spatial compression in visual neglect: A case
study. Cortex 27:623-629.
Halligan, P. W.,
and J. C. Marshall. (1994). Figural perception and parsing in visuospatial
neglect. Neuroreport 5:537-539.
Heilman, K. M., R.
T. Watson, and E. Valenstein. (1985). Neglect and related disorders. In K. M.
Heilman and E. Valenstein, Eds., Clinical Neuropsychology. New
York: Oxford University Press, pp. 243-294.
Karnath, H. O., P.
Schenkel, and B. Fischer. (1991). Trunk orientation as the determining factor
of the "contralateral" deficit in the neglect syndrome and as the
physical anchor of the internal representation of body orientation in space. Brain
114:1997-2014.
Kinsbourne, M.
(1987). Mechanisms of unilateral neglect. In M. Jeannerod, Ed., Neurophysiological
and Neuropsychological Aspects of Spatial Neglect. Amsterdam:
North-Holland, pp. 69-86.
Ladavas, E.
(1987). Is the hemispatial deficit produced by right parietal lobe damage
associated with retinal or gravitational coordinates? Brain
110:167-180.
Ladavas, E., M. D.
Pesce, and L. Provinciali. (1989). Unilateral attention deficits and
hemispheric asymmetries in the control of visual attention. Neuropsychologia
27(3):353-366.
Mattingley, J. B.,
G. Davis, and J. Driver. (1997). Preattentive filling-in of visual surfaces in
parietal extinction. Science 275:671-674.
McGlinchey-Berroth,
R, W. P. Milberg, M. Verfaellie, L. Grande, M. D'Esposito, and M. Alexandre.
(1996). Semantic processing and orthographic specificity in hemispatial
neglect. Journal of Cognitive Neuroscience 8:291-304.
Mozer, M. C., and
M. Behrmann. (1990). On the interaction of selective attention and lexical
knowledge: A connectionist account of neglect dyslexia. Journal of
Cognitive Neuroscience 2(2):96-123.
Mozer, M. C., P.
W. Halligan, and J. C. Marshall. (1997). The end of the line for a
brain-damaged model of hemispatial neglect. Journal of Cognitive
Neuroscience 9(2):171-190.
Posner, M. I., J.
A. Walker, F. J. Friedrich, and R. D. Rafal. (1984). Effects of parietal injury
on covert orienting of visual attention. Journal of Neuroscience
4:1863-1877.
Pouget, A., and T.
J. Sejnowski. (1997a). Lesion in a basis function model of spatial
representations: Comparison with hemineglect. In P. Thier and H. O. Karnath,
Eds., Parietal Lobe Contribution in Orientation in 3D Space.
Springer.
Pouget, A., and T.
J. Sejnowski. (1997b). Spatial transformations in the parietal cortex using
basis functions. Journal of Cognitive Neuroscience 9(2):222-237.
Rizzolatti, G.,
and A. Berti. (1990). Neglect as a neural representation deficit. Revue
Neurologique 146(10):626-634.
Tipper, S. P., and
M. Behrmann. (1996). Object-centered not scene-based visual neglect. Journal
of Experimental Psychol ogy, Human Perception and Performance
22(5):1261-1278.
![]() Copyright © 1999 Massachusetts
Institute of Technology
Copyright © 1999 Massachusetts
Institute of Technology